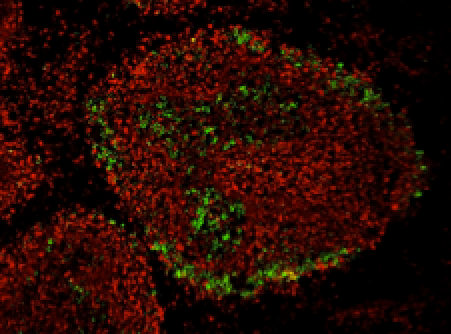
Discrete virus factories form in the cytoplasm of cells co-infected with two replication competent tagged reporter birnaviruses, that subsequently coalesce over time
The Birnaviridae family, responsible for major economic losses to poultry and aquaculture, are non-enveloped viruses with a segmented double-stranded (ds)RNA genome that replicate in discrete cytoplasmic virus factories (VFs). Reassortment is common, however, the underlying mechanism remains unknown given that VFs may act as a barrier to genome mixing. In order to provide new information on VF trafficking during dsRNA virus co-infection, we rescued two recombinant infectious bursal disease viruses (IBDVs) of strain PBG98 containing either a split GFP11- or Tetracysteine (TC)- tag fused to the VP1 polymerase (PBG98-VP1-GFP11 and PBG98-VP1-TC). DF-1 cells transfected with GFP1-10 prior to PBG98-VP1-GFP11 infection, or stained with ReAsH following PBG98-VP1-TC infection, had green or red foci in the cytoplasm respectively that co-localised with VP3 and dsRNA, consistent with VFs. The average number of VFs decreased from a mean of 60 to 5 per cell between 10 and 24 hours post infection (hpi) (p<0.0001), while the average area increased from 1.24 μm(2) to 45.01μm(2) (p<0.0001), and live cell imaging revealed that the VFs were highly dynamic structures that coalesced in the cytoplasm. Small VFs moved faster than large (average 0.57μm/s at 16 hpi compared to 0.22 μm/s at 22 hpi), and VF coalescence was dependent on an intact microtubule network and actin cytoskeleton. During co-infection with PBG98-VP1-GFP11 and PBG98-VP1-TC viruses, discrete VFs initially formed from each input virus that subsequently coalesced 10-16 hpi, and we speculate that Birnaviridae reassortment requires VF coalescence.
IMPORTANCE Reassortment is common in viruses with segmented double stranded (ds)RNA genomes. However, these viruses typically replicate within discrete cytoplasmic virus factories (VFs) that may represent a barrier to genome mixing. We generated the first replication competent tagged reporter birnaviruses, infectious bursal disease viruses (IBDVs) containing a split GFP11 or tetracysteine (TC) tag and used the viruses to track the location and movement of IBDV VFs, in order to better understand the intracellular dynamics of VFs during a co-infection. Discrete VFs initially formed from each virus that subsequently coalesced from 10 hours pi. We hypothesise that VF coalescence is required for the reassortment of the Birnaviridae. This study provides new information that adds to our understanding of dsRNA virus VF trafficking.